

摘要
本发明提供一种产苹果酸基因工程菌及其生产苹果酸的方法,包括提高受体菌中二元酸转 运蛋白的表达或活性,降低受体菌中甲基转移酶 laeA的表达或活性,获得产苹果酸的工程菌。本发明有效提高苹果酸的产量,进一步地,通过引入耐酸工程菌,还有效避免中和剂的使用。
权利要求书
1.一种构建产苹果酸的工程菌的方法,其特征在于:包括提高受体菌中二元酸转运蛋白的表达或活性,降低所述受体菌中甲基转移酶laeA的表达或活性,获得产苹果酸的工程菌。
2.根据权利要求1所述的方法,其特征在于:所述二元酸转运蛋白的氨基酸序列包括如 SEQ ID NO.1所示的氨基酸序列或保有功能的随机和/或定点诱变序列;
和/或,提高所述受体菌中二元酸转运蛋白的表达或活性是通过向受体菌中导入二元 酸转运蛋白的编码基因来实现的;
和/或,所述二元酸转运蛋白的编码基因的核苷酸序列包括如SEQ ID NO .2所示的片段 或保有功能的随机和/或定点诱变序列。
3.根据权利要求1所述的方法,其特征在于:所述甲基转移酶laeA的氨基酸序列包括如 SEQ ID NO.3所示的氨基酸序列或保有功能的随机和/或定点诱变序列。
和/或,降低受体菌中所述甲基转移酶laeA的表达或活性是通过敲除或沉默所述甲基 转移酶laeA的编码基因来实现;
和/或,所述甲基转移酶laeA的编码基因的核苷酸序列包括如SEQ ID NO .4所示的片段 或保有功能的随机和/或定点诱变序列。
4.根 据 权 利 要 求 1 所 述 的 方 法 ,其 特 征 在 于 :所 述 受 体 菌 选 自 毁 丝 霉 属 (Myceliophthora)、曲霉属(Aspergillus)中的至少一种,优选地,所述毁丝霉属菌株选自 嗜 热 毁 丝 霉 ( M y c e l i o p h t h o r a t h e r m o p h i l a ) 、异 梭 毁 丝 霉 (Myce l iophthoraheterotha l l ica) 中 的至少一种,所述曲霉属菌株选自黑曲霉 (Aspergillus niger)、米曲霉(Aspergillus oryzae)、土曲霉(Aspergillus terreus)、酱 油曲霉(Aspergillus sojae)中的至少一种。
5.根据权利要求1所述的方法,其特征在于:通过同源重组法、原生质体法、农杆菌介导 法、RNAi沉默法或CRISPR法降低所述甲基转移酶laeA的表达或活性。
6.根据权利要求1所述的方法,其特征在于:所述受体菌选自嗜热毁丝霉,并采用同源 重组法敲除所述甲基转移酶laeA的编码基因,优选地,进行敲除时,上游同源重组片段的核 苷酸序列包括如SEQ ID NO.5所示片段或保有功能的随机和/或定点诱变序列,下游同源重 组片段的核苷酸序列包括如SEQ ID NO.6所示片段或保有功能的随机和/或定点诱变序列。
7.根据权利要求1所述的方法,其特征在于:所述受体菌选自黑曲霉时,并采用同源重 组法敲除所述甲基转移酶laeA的编码基因,优选地,进行敲除时,上游同源重组片段的核苷 酸序列包括如SEQ ID NO .7所示片段或保有功能的随机和/或定点诱变序列,下游同源重组 片段的核苷酸序列包括如SEQ ID NO.8所示片段或保有功能的随机和/或定点诱变序列。
8.根据权利要求1-7任意一项所述方法制得的用于生产苹果酸的工程菌。 9.根据权利要求8所述工程菌在苹果酸生产中的应用。
10.一种生产苹果酸的方法,包括:培养权利要求8所述工程菌,收集孢子,将所述孢子 接种至发酵培养基中,收集发酵产物,从中获得苹果酸,优选地,所述发酵培养基包括如下 组分:100g/L葡萄糖,6 .0g/L蛋白胨,150mg/L磷酸二氢钾,150mg/L磷酸氢二钾,100mg/L氯 化钙,100mg/L硫酸镁,碳酸钙80.0g/L,1mg/L生物素,1mL/L微量元素,余量为水;其中,所述 微量元素包括如下组分:氯化钠5g/L,硫酸亚铁5g/L,柠檬酸1g/L,余量为水。
说明书
技术领域
[0001] 本发明涉及基因工程和生物技术领域,特别是涉及一种通过基因工程手段构建苹 果酸生产菌株和提高苹果酸产量的方法及应用,同时还进一步涉及耐酸的高产苹果酸生产 菌株的构建及应用。
背景技术
[0002] 生物法生产大宗化学品是我国经济绿色可持续发展、摆脱石油依赖的有效途径之 一。苹果酸是非常重要的大宗化工产品,早在2004年就被美国能源部认定为最具竞争力的 12种平台化合物之一,其被广泛用于食品、饮料、香料、医药保健、化工、塑料等行业。目前, 国际市场需求量保持快速增长,市场前景广阔,实现发酵法生产苹果酸对我国建立化学品 生物制造产业体系具有重要意义。
[0003] 目前,合成苹果酸的相关研究主要集中在丝状真菌上。早在上世纪60年代,就有利 用丝状真菌黄曲霉合成苹果酸的报道。然而,由于黄曲霉毒素的原因,利用黄曲霉发酵生产 苹果酸未能得到大规模工业生产。近年来,有报道表明在米曲霉A.oryzae NRRL 3488过表 达自身的丙酮酸脱氢酶、苹果酸脱氢酶及C4-二羧酸转运蛋白后,其苹果酸产量得到了极大 的提高,以葡萄糖为碳源时,在164小时内苹果酸产量达到了154g/L,产率为0 .94g/L/h,转 化率达到理论值的69%。国内江南大学对米曲霉苹果酸合成途径进行了系统代谢工程改 造,苹果酸产量达到93 .2g/L,生产效率为1 .38g/L/h。在苹果酸合成过程中,中和剂碳酸钙 与苹果酸反应,将培养基的pH保持在中性左右,利于微生物发酵生长,但会增加发酵成本, 同时形成的苹果酸钙在后期的分离纯化中需要用到大量强酸,不仅增加了成本,还对环境 造成了极大的危害。因此,开发耐酸的苹果酸生产菌株是解决这一问题的有效途径。
[0004] 曲霉被广泛的应用在蛋白、化学品的工业化生产过程中,尤其是黑曲霉,它是多种 蛋白和有机酸的生产菌株。黑曲霉在深层次发酵过程中具有极其显著的特点,在快速生长 期时,其可以分泌大量有机酸(草酸、柠檬酸等),使得发酵培养基的pH低于2。黑曲霉的这种 发酵特性,不仅大大降低发酵过程中染菌的风险,还可以省去生产有机酸过程中中和有机 酸所使用的大量碳酸盐,对环境友好。然而,目前黑曲霉主要还是集中在柠檬酸的生产上, 并没有涉及苹果酸的生产。
发明内容
[0005] 鉴于以上所述现有技术的缺点,本发明的目的在于提供一种产苹果酸基因工程菌 及其生产苹果酸的方法, 用于提高苹果酸产量,在此基础上进一步提高工程菌的耐酸性,避 免中和剂的使用。
[0006] 为实现上述目的及其他相关目的,本发明提供一种构建产苹果酸的工程菌的方 法,包括提高受体菌中二元酸转运蛋白的表达或活性,降低所述受体菌中甲基转移酶laeA 的表达或活性,获得产苹果酸的工程菌。
[0007] 可选地,所述二元酸转运蛋白的氨基酸序列包括如SEQ ID NO .1所示的氨基酸序列或保有功能的随机和/或定点诱变序列。
[0008] 可选地,提高上述受体菌中二元酸转运蛋白的表达或活性是通过向受体菌中导入 二元酸转运蛋白的编码基因来实现的。
[0009] 可选地,所述二元酸转运蛋白的编码基因的核苷酸序列包括如SEQ ID NO .2所示 的片段或保有功能的随机和/或定点诱变序列。
[0010] 可选地,所述甲基转移酶laeA的氨基酸序列包括如SEQ ID NO .3所示的氨基酸序 列或保有功能的随机和/或定点诱变序列。
[0011] 可选地,降低所述受体菌中甲基转移酶laeA的表达或活性是通过敲除或沉默所述 受体菌中甲基转移酶laeA的编码基因来实现。
[0012] 可选地,所述甲基转移酶laeA的编码基因的核苷酸序列包括如SEQ ID NO .4所示 的片段或保有功能的随机和/或定点诱变序列。
[0013] 可选地,所述受体菌选自毁丝霉属(Myceliophthora)、曲霉属(Aspergillus)中的 至少一种。
[0014] 可选地,所述毁丝霉属菌株选自嗜热毁丝霉(Myceliophthora thermophila)、异 梭毁丝霉(Myceliophthoraheterothallica)中的至少一种。
[0015] 可选地,所述曲霉属菌株选自黑曲霉(Aspergillus niger)、米曲霉(Aspergillus oryzae)、土曲霉(Aspergillus terreus)、酱油曲霉(Aspergillus sojae)中的至少一种。 [0016] 可选地,通过同源重组法、原生质体法、农杆菌介导法、RNAi沉默法或CRISPR法降 低所述甲基转移酶laeA的表达或活性。
[0017] 可选地,所述受体菌选自嗜热毁丝霉,并采用同源重组法敲除所述甲基转移酶 laeA的编码基因。
[0018] 可选地,进行敲除时,上游同源重组片段的核苷酸序列包括SEQ ID NO .5所示片段 或保有功能的随机和/或定点诱变序列,下游同源重组片段的核苷酸序列包括SEQ ID NO .6 所示片段或保有功能的随机和/或定点诱变序列。
[0019] 可选地,所述受体菌选自黑曲霉,并采用同源重组法敲除所述甲基转移酶laeA的 编码基因。
[0020] 可选地,其上游同源重组片段的核苷酸序列包括SEQ ID NO .7所示片段或保有功 能的随机和/或定点诱变序列,下游同源重组片段的核苷酸序列包括SEQ ID NO .8所示片段 或保有功能的随机和/或定点诱变序列。
[0021] 由上述方法制备得到的用于生产苹果酸的工程菌也属于本发明的保护范围。
[0022] 上述工程菌在苹果酸生产中的应用也属于本发明的保护范围。
[0023] 本发明还提供生产苹果酸的方法,包括:培养所述工程菌,收集孢子,将所述孢子接种至发酵培养基中,收集发酵产物,从中获得苹果酸。
[0024] 可选地,所述发酵培养基包括如下组分:100g/L葡萄糖,6.0g/L蛋白胨,150mg/L磷 酸二氢钾,150mg/L磷酸氢二钾,100mg/L氯化钙,100mg/L硫酸镁,碳酸钙80 .0g/L,1mg/L生 物素,1mL/L微量元素,余量为水。
[0025] 可选地,所述微量元素包括如下组分:氯化钠5g/L,硫酸亚铁5g/L,柠檬酸1g/L,余 量为水。
[0026] 如上所述,本发明的产苹果酸基因工程菌及其生产苹果酸的方法,具有以下有益效果:本发明寻找到有效提高苹果酸产量的新途径,具体是通过降低受体菌中甲基转移酶laeA的表达或活性,并提高受体菌中二元酸转运蛋白的表达或活性来实现,进一步地,本发 明提供了一种耐酸苹果酸工程菌及其生产苹果酸的方法,包括降低受体菌中甲基转移酶 laeA的表达或活性,并提高受体菌中二元酸转运蛋白的表达或活性,获得具有耐酸特性的 苹果酸工程菌株,避免了中和剂的使用。
附图说明
[0027] 图1显示为二元酸转运蛋白mae基因表达载体pGL01-mae的质粒图谱。
[0028] 图2显示为嗜热毁丝霉野生型菌株及其突变株的苹果酸产量图。
[0029] 图3显示为敲除嗜热毁丝霉laeA的敲除载体pKO-laeA的质粒图谱。
[0030] 图4显示为敲除黑曲霉laeA的敲除载体pGL01-mae-KOlaeA的质粒图谱。
[0031] 图5显示为苹果酸产量随时间的变化曲线图。
[0032] 图6显示为黑曲霉野生型及其突变株的柠檬酸和苹果酸产量图。
具体实施方式
[0033] 以下通过特定的具体实例说明本发明的实施方式,本领域技术人员可由本说明书 所揭露的内容轻易地了解本发明的其他优点与功效。本发明还可以通过另外不同的具体实 施方式加以实施或应用,本说明书中的各项细节也可以基于不同观点与应用,在没有背离 本发明的精神下进行各种修饰或改变。
[0034] 在丝状真菌中,甲基转移酶可以调控次生代谢物合成过程中相关基因的表达水 平,进而影响次生代谢物的积累水平。在黑曲霉中敲除或下调甲基转移酶可以减少柠檬酸 的生产,本发明意外地发现,敲除黑曲霉的甲基转移酶的同时引入二元酸转运蛋白,可以获 得耐酸的苹果酸生产菌株,这种新方法未见文献、专利报道。并且,本发明还意外地发现,敲 除嗜热毁丝霉中的一个甲基转移酶,可以提高苹果酸的产量,目前尚未有关于甲基转移酶 调控苹果酸的合成途径的报道,其作用原理也尚不清楚,不能由已知报道推测获得。
[0035] 以下实施例采用的察氏固体培养基组成如下:NaNO3 3g/L,K2HPO4 1g/L,MgSO4 · 7H2O0.5g/L,KCl 0.5g/L,FeSO4 0.01g/L,蔗糖30g/L,琼脂粉20g/L,余量为水,总体积为1L。 [0036] 下述实施例中涉及分子生物学操作所用的限制性内切酶均购于NEB公司,质粒提 取和DNA回收试剂盒均购于天根生化科技(北京)有限公司。
[0037] 下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
[0038] 下述实施例中所使用的材料、试剂,如无特殊说明,均可从商业途径购得。
[0039] 下述实施例中,质粒pGL01的核苷酸序列如SEQ ID NO .11所示,质粒pKO的核苷酸序列如SEQ ID NO.12所示。
[0040] 实施例1
[0041] 在嗜热毁丝霉(Myceliophthora thermophila)中过表达mae基因
[0042] 1.过表达二元酸转运蛋白载体构建及转化
[0043] 利用引物Mae-F(SEQ ID NO .13:5'-aatggttaacATGCTGACACCTCCCAAG-3’)和引物Mae-R(SEQ ID NO.14:5'-aatggttaacCTAATCAGATACATCCTC-3'),从米曲霉cDNA中扩增二元 酸转运蛋白基因(简称mae基因)的编码阅读框,PCR反应体系为:2×Rapid Taq Master Mix25μL,Mae-F 1μL,Mae-R 1μL,cDNA1μL,水22μL。PCR反应条件为:95℃3min;95℃15s,58℃15s,72℃1 .5min,30个循环;72℃5min,4℃5min。PCR反应结束后,使用限制性内切酶Hpa I 消化PCR产物,并将其插入到质粒pGL01的EcoR V位点 (质粒图谱如图1所示),得到质粒 pGL01-mae。
[0044] 2.嗜热毁丝霉原生质体法转化方法
[0045] 原生质体转化方法参考山东大学张美玲硕士论文“里氏木霉信号传导途径对纤维 素酶的合成调控”,具体如下:嗜热毁丝霉(美国典型菌种保藏中心,ATCC424)在察氏固体培 养基上在45℃条件下生长1周左右, 用生理盐水收集孢子,获得新鲜孢子悬液。在察氏固体 培养基上铺一层玻璃纸,加100μl孢子悬液,并用涂布棒轻轻涂匀,以防止气泡产生,大约做 10个平板,45℃培养12-16小时。称取0.2g裂解酶(购自Sigma-Aldrich公司,货号L1412),溶 于30mL溶液I(山梨醇1 .2M,磷酸二氢钾0 .1M,pH5 .5)中,混匀,过滤除菌,获得裂解液。吸取 适量裂解液于无菌的培养皿中,将长有菌丝的玻璃纸倒扣在培养皿中,再加入适量裂解液, 然后再放入第二张长有菌丝的玻璃纸,以此类推,直到玻璃纸全放进去,30℃静置裂解1 .5- 2小时。裂解后, 用无菌镊子取出玻璃纸,并用3层擦镜纸过滤,获得含有原生质体的滤液,4 ℃条件下2000rpm离心15分钟,弃上清, 用4mL溶液II(山梨醇1 .0M,Tris-HCl10mM,CaCl2 50mM,pH7.5)重悬原生质体,4℃条件下2000rpm离心15分钟,弃上清。用200μL溶液II将原生 质体重悬,待用。将200μL原生质体、10μg转化DNA片段、50μL溶液III(PEG6000 25%,CaCl2 50mM,Tris-HCl 10mM,pH7 .5)混匀,冰上放置20分钟,然后加入2mL溶液III,室温静置5分 钟,最后加入4mL溶液II,混匀。将上述混合物与50℃的含有相应抗生素的上层培养基混匀 后,迅速倒在凝固的含有相应抗生素的下层平板中,凝固后45℃培养,待转化子长出后,挑 至新鲜的含有相应抗生素的察氏培养基中生长。转化中所用下层培养基为山梨醇182g/L、 NaNO3 3g/L、K2HPO4 1g/L、MgSO4 ·7H2O 0.5g/L、KCl 0.5g/L、FeSO4 0.01g/L、蔗糖30g/L,琼 脂20g/L,加水定容至1L,121℃灭菌15min;上层培养基为:山梨醇182g/L,NaNO3 3g/L, K2HPO4 1g/L,MgSO4 ·7H2O 0.5g/L,KCl 0.5g/L,FeSO4 0.01g/L,蔗糖30g/L,琼脂糖20g/L, 余量为水,加水定容至1L,121℃灭菌15min。
[0046] 将质粒pGL01-mae按上述方法转入嗜热毁丝霉中,将转化子挑取至含有80μg/mL的 潮霉素的察氏培养基中。
[0047] 3.丝状真菌基因组提取
[0048] 挑取少量菌丝到含有石英砂的300μL抽提液(200mM Tris,250mMNaCl,25mMEDTA, 2%SDS,pH8.5)中,旋涡剧烈振荡10min,65℃水浴30min;加入300μL苯酚/氯仿/异戊醇混合 液(按体积计,苯酚:氯仿:异戊醇=25:24:1)后旋涡混匀30s,12000rpm室温离心10min,收 集上清至另一离心管中;加入2 .5倍上清液体积的无水乙醇,混匀,-20℃放置2小时; 12000rpm室温离心10min,弃上清,用70%乙醇 (5000rpm,5min)洗涤两次,4℃条件下 12000rpm离心10min,弃上清,待乙醇挥发后,加入50μL无菌水,溶解基因组,待用。
[0049] 4.重组真菌转化子MTmae生产苹果酸能力表征
[0050] 将转化获得的转化子按上述方法提取基因组作为模板, 用引物Mae-F和Mae-R进行 扩增,获得相应条带的转化子即为正确转化子,命名为MTmae,待后续发酵。将转化子MTmae 接种于察氏固体培养基上,45℃培养14天,收集孢子,将上述孢子以1×106个/mL的接种量, 接种至50mL发酵培养基中(发酵培养基组成如下:100g葡萄糖,6.0g蛋白胨,150mg磷酸二氢钾,150mg磷酸氢二钾,100mg氯化钙,100mg硫酸镁,碳酸钙80.0g,1mg生物素,1mL微量元素,余量为水,总体积为1L。其中,微量元素组成如下:氯化钠5g,硫酸亚铁5g,柠檬酸1g,余量为 水,总体积为1L),45℃,120rpm培养,第五天取样测定苹果酸含量,经过HPLC分析,图2显示 为嗜热毁丝霉野生型菌株及其突变株的苹果酸产量图,MTmae的苹果酸产量为50 .8g/L,即 每升培养基中所产生的苹果酸为50.8g,而出发菌株几乎不产苹果酸。
[0051] 实施例2
[0052] 在嗜热毁丝霉重组菌中敲除甲基转移酶laeA基因
[0053] 1.构建嗜热毁丝霉甲基转移酶laeA基因敲除载体
[0054] 根据嗜热毁丝霉全基因组信息,设计引物扩增得到甲基转移酶laeA基因(MYCTH_2294559)上下游同源片段,引物设计如下所示:
[0055] laeA-MFF(SEQ ID NO.15):TTCGAGATCTAACCCATCATCTCGGTCCTCTA;
[0056] laeA-MFR(SEQ ID NO.16):TTCGAGATCTAGCTTCAAATCGCGTGTTTCAT;
[0057] laeA-MRF(SEQ ID NO.17):TTCGACTAGTGGCAAGCCCTCCAGCCACTT;
[0058] laeA-MRR(SEQ ID NO.18):TTCGACTAGTTTCCTGGGCAGCAGCGGAGT;
[0059] laeA-F(SEQ ID NO.19):AATCGCACCTACCAAGAAGT;
[0060] laeA-R(SEQ ID NO.20):ATGGTCAGTCAGCAGTGTTT。
[0061] 以基因组为模板,进行PCR扩增,用引物laeA-MFF和laeA-MFR扩增获得上游同源片段, 用引物laeA-MRF和laeA-MRR扩增获得下游同源片段,将上下游同源片段分别插入敲除 质粒pKO的Bgl II和SpeI位点,如图3所示,获得质粒pKO-laeA。以质粒pKO-laeA为模板, 用 引物laeA-MFF和laeA-MRR扩增得到敲除片段,参照实施例1中的原生质体转化方法,将其转 入重组子MTmae中,将转化子挑至含有100μg/mL G418的察氏培养基中,提取基因组, 用引物 laeA-F和laeA-R进行验证,扩增不出条带的为正确转化子,命名为MTmaeΔlaeA。
[0062] Mt上游同源臂核苷酸序列如SEQ ID NO.5所示。
[0063] Mt下游同源臂核苷酸序列如SEQ ID NO.6所示。
[0064] Mt laeA同源臂核苷酸序列如SEQ ID NO .9所示,其氨基酸序列如SEQ ID NO .10所示。
[0065] 2.甲基转移酶敲除菌株MTmaeΔlaeA产苹果酸能力表征
[0066] 将转化子MTmaeΔlaeA接种于察氏固体培养基上,45℃培养14天,收集孢子,将上述孢子以1×106个/mL的接种量,接种至50mL发酵培养基(与实施例1中发酵培养基相同), 45℃,120rpm培养,第五天取样测定苹果酸含量,经过HPLC分析,如图2所示,MTmaeΔlaeA的 苹果酸产量为75g/L,与对照菌株MTmae相比,苹果酸产量提高了50%。
[0067] 如图5所示为苹果酸产量随时间的变化情况图,从图中可以看出,随时间的延长, 菌株Mtmae、菌株MtmaeΔlaeA的苹果酸产量持续升高,到第五天最高,后续变化较小,并且, 我们意外地发现,菌株Mtmae的苹果酸产量显著低于菌株MtmaeΔlaeA的苹果酸产量。
[0068] 实施例3
[0069] 在黑曲霉(Aspergillus niger)中敲除甲基转移酶laeA同时过表达二元酸转运蛋白
[0070] 1.构建黑曲霉甲基转移酶AnlaeA基因敲除载体
[0071] 根据黑曲霉(荷兰微生物菌种保存中心,CBS513.88)的全基因组信息,设计引物扩增得到甲基转移酶laeA基因(Aspni7︱170198)上下游同源片段,引物设计如下所示:
[0072] laeA-AFF(SEQ ID NO.21):ttcgagatctTTTCCCACCATTCCAGCCCT;
[0073] laeA-AFR(SEQ ID NO.22):ttcgagatctATCGTTGTTGTCCGGGGGTT;
[0074] laeA-ARF(SEQ ID NO.23):ttcgactagtTGAGCAGAAGGTAGCCCGTT;
[0075] laeA-ARR(SEQ ID NO.24):ttcgactagtGAGGGTTCTTGGCGTCTGGT。
[0076] 以基因组为模板,进行PCR扩增,用引物laeA-AFF和laeA-AFR扩增获得上游同源片段, 用引物laeA-ARF和laeA-ARR扩增获得下游同源片段,PCR方法参照实施例1,As上游同源 臂核苷酸序列如序列7所示,As下游同源臂核苷酸序列如序列8所示,将上下游同源片段分 别用BglII和SpeI消化,再将消化后的上下游片段插入质粒pGL01-mae相应的酶切位点中, 获得质粒pGL01-mae-KOlaeA,其物理图谱如图4所示。
[0077] 2.黑曲霉原生质体转化
[0078] 黑曲霉在察氏培养基上30℃生长14天, 用生理盐水洗下孢子, 制成孢子悬液。按1 ×106个/mL的浓度接种孢子至ME液体培养基 (麦芽提取物30g/L,蛋白胨5g/L,121℃ , 15min)中,30℃条件下200rpm培养过夜。用三层擦镜滤纸收集菌体,将菌体转至30mL裂解液 中,35℃条件下100rpm振荡培养1 .5-3小时,4℃条件下2000rpm离心10min,弃上清,加入相 同体积的预冷的STC(1 .2M山梨醇,50mMCaCl2,10mM Tris,pH 7.5,121℃,15min),混匀,4℃ 条件下2000rpm离心10min,弃上清,加入100μLSTC(25%PEG6000,50mM CaCl2,10mM Tris, pH 7.5,121℃,15min),混匀,得到原生质体悬液,待用。
[0079] 上述裂解液的配制方法如下:称取0 .2g裂解酶 (购自Sigma-Aldrich公司, 货号 L1412),溶于30mL MC溶液,过滤灭菌得到。其中,MC溶液的组成如下:50mM CaCl2,20mM MES/NaOH,pH 5 .8,渗透压稳定剂(0 .9M KCl或1 .2M山梨醇或0 .4M MgSO4),121℃灭菌 15min。
[0080] 100μL原生质体加入10μgDNA片段和330μLPEG缓冲液,冰上放置20min,加入 2mLPEG,室温放置10min,加入4mLSTC,混匀,再加入4mL于50℃预热的上层培养基,混匀后倒 在含有100μg/mL潮霉素的下层培养基上,平板在30℃培养直至出现菌落。
[0081] 以步骤2制得的质粒pGL01-mae-KOlaeA为模板, 用引物laeA-AFF和laeA-ARR扩增 得到敲除片段, 用上述原生质体法将DNA片段转入黑曲霉中,提取转化子基因组,利用PCR进 行检测。用引物Mae-F和Mae-R可以扩增获得条带且用引物laeA-AnF和laeA-AnR不能扩增获 得条带的转化子为正确转化子,命名为ANmae。
[0082] laeA-AnF(SEQ ID NO.25):ACAACTACCGGGCGACTAAT;
[0083] laeA-AnR(SEQ ID NO.26):GATCTGGGTAAAGCCTGCCT。
[0084] 3.黑曲霉重组转化子ANmae产苹果酸能力表征
[0085] 将转化子ANmae接种于察氏固体培养基上,30℃培养14天,收集孢子,将上述孢子以1×106个/mL的接种量,接种至50mL柠檬酸发酵培养基中(按质量占比计,柠檬酸发酵培 养基配方如下:蔗糖10% ,硝酸铵0 .25% ,硫酸镁0 .1% ,磷酸二氢钾0 .1% ,酵母浸出粉 0 .05%,余量为水,总体积为1L,pH 2.5,调pH所用试剂为盐酸),30℃,250rpm培养,第五天 取样测定苹果酸含量;同时,将黑曲霉野生型菌株接种至上述柠檬酸发酵培养基中,作为对 照组,经过HPLC分析可知,如图6所示,黑曲霉野生型菌株的柠檬酸产量为43g/L,而几乎不 产苹果酸。相反地,几乎检测不到转化子ANmae的柠檬酸产量,而苹果酸产量为35g/L。因此,敲除甲基转移酶laeA的黑曲霉是极佳的生产苹果酸的真菌底盘。
[0086] 综上所述,甲基转移酶laeA参与丝状真菌次生代谢物和初级代谢物的合成,敲除 甲基转移酶LaeA的编码基因可以提高某些代谢物的产量,也会降低某些代谢物的产量,因 此该基因在黑曲霉或嗜热毁丝霉等受体菌中对于苹果酸合成的影响是无法推测的,需要具 体试验探索。将两者结合起来,创建新型耐酸苹果酸生产细胞工厂,对苹果酸的工业化生产 具有重要意义。
[0087] 曲霉是有机酸发酵的传统优势菌株,尤其是黑曲霉的柠檬酸发酵,黑曲霉具有极 好的耐酸性,不但在发酵过程中不易污染,而且生产有机酸过程中不需要添加中和剂(如碳 酸钙等碳酸盐),因此,该生产工艺对环境友好且具有较低的生产成本。而目前苹果酸的发 酵生产都依赖于中和剂的使用,因此,构建不依赖中和剂的黑曲霉-苹果酸细胞工厂是具有 创新性的,无法由现有文献数据推测获得,需要具体试验探索。
[0088] 上述实施例仅例示性说明本发明的原理及其功效,而非用于限制本发明。任何熟 悉此技术的人士皆可在不违背本发明的精神及范畴下,对上述实施例进行修饰或改变。因 此,举凡所属技术领域中具有通常知识者在未脱离本发明所揭示的精神与技术思想下所完 成的一切等效修饰或改变,仍应由本发明的权利要求所涵盖。
序列表
SEQUENCE LISTING
<110>上海东庚化工技术有 限公司
<120> 一种产苹果酸基因工程菌及其生产苹果酸的方法
<130> PCNDG192395
<160> 26
<170> PatentIn version 3. 5
<210> 1
<211> 380
<212> PRT
<213> Artifici al
<220>
<223>二元酸转运蛋白mae的氨基酸序列
<400> 1

<210> 2
<211> 1143
<212> DNA
<213> Artificial
<220>
<223> 二元酸转运蛋白mae的核苷酸序列
<400> 2

<210> 3
<211> 375
<212> PRT
<213> Artificial
<220>
<223> 甲基转移酶laeA的氨基酸序列
<400> 3

<210> 4
<211> 1128
<212> DNA
<213> Artificial
<220>
<223> 甲基转移酶laeA的核苷酸序列
<400> 4

<210> 5
<211> 995
<<212> DNA
<213>Artificial
<220>
<223> Mt上游同源臂
<400> 5

<210> 6
<211> 958
<212> DNA
<213> Artificial
<220>
<223> Mt下游同源臂
<400> 6

<210> 7
<211> 1216
<212> DNA
<213> Artificial
<220>
<223> As上游同源臂
<400> 7

<210> 8
<211> 1099
<212> DNA
<213> Artificial
<220>
<223> As下游同源臂
<400> 8

<210> 9
<211> 969
<212> DNA
<213> Artificial
<220>
<223> Mt laeA同源臂核苷酸序列
<400> 9

<210> 10
<211> 322
<212> PRT
<213> Artificial
<220>
<223> Mt laeA同源臂氨基酸序列
<400> 10
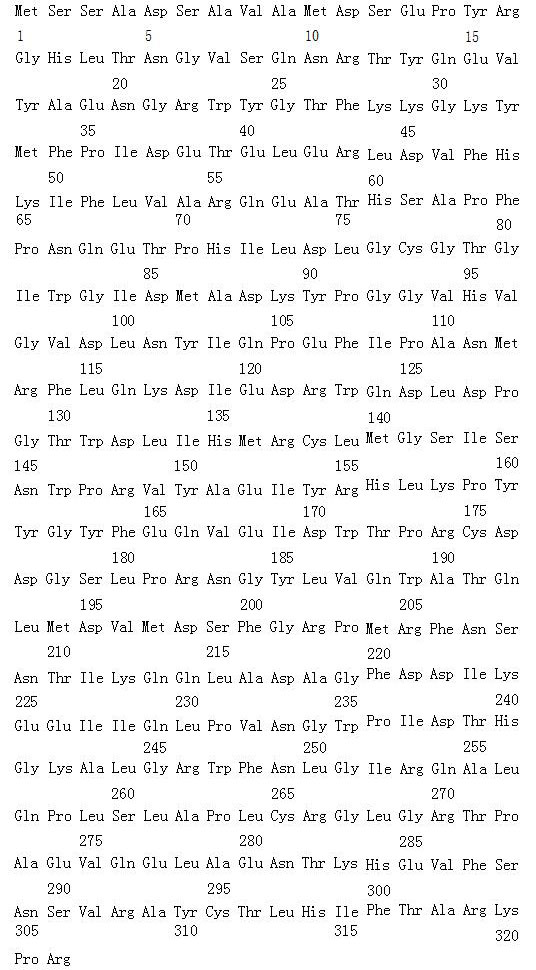
<210> 11
<211> 5475
<212> DNA
<213> Artificial
<220>
<223> pGL01
<400> 11

<210> 12
<211> 3828
<212> DNA
<213> Artificial
<220>
<223>pK0
<400>12

<210> 13
<211> 28
<212> DNA
<213> Artificial
<220>
<223>引物Mae-F
< 400>13
aatggttaac atgctgacac ctcccaag 28
<210> 14
<211>28
<212> DNA
<213> Artificial
<220>
<223>引物Mae-R
<400>14
aatggttaac ctaatcagat acatcctc 28
<210>15
<211>32
<212>DNA
<213>Artificial
<220>
<223>laeA- -MFF
<400>15
ttcgagatct aacccatcat ctcggtcctc ta 32
<210> 16
<211> 32
<212> DNA
<213> Artificial
<220>
<223> laeA-MFR
<400>16
ttcgagatct agcttcaaat cgcgtgtttc at 32
<210> 17
<211> 30
<212> DNA
<213> Artificial
<220>
<223> laeA-MRF
<400> 17
ttcgactagt ggcaagccct ccagccactt 30
<210> 18
<211> 30
<212> DNA
<213> Artificial
<220>
<223> laeA-MRR
<400> 18
ttcgactagt ttcctgggca gcagcggagt 30
<210> 19
<211> 20
<212> DNA
<213> Artificial
<220>
<223> laeA-F
<400> 19
aatcgcacct accaagaagt 20
<210> 20
<211> 20
<212> DNA
<213> Artificial
<220>
<223> laeA-R
<400> 20
atggtcagtc agcagtgttt 20
<210> 21
<211> 30
<212> DNA
<213> Artificial
<220>
<223> laeA-AFF
<400> 21
ttcgagatct tttcccacca ttccagccct 30
<210> 22
<211> 30
<212> DNA
<213> Artificial
<220>
<223> laeA-AFR
<400> 22
ttcgagatct atcgttgttg tccgggggtt 30
<210> 23
<211> 30
<212> DNA
<213> Artificial
<220>
<223> laeA-ARF
<400> 23
ttcgactagt tgagcagaag gtagcccgtt 30
<210> 24
<211> 30
<212> DNA
<213> Artificial
<220>
<223> laeA-ARR
<400> 24
ttcgactagt gagggttctt ggcgtctggt 30
<210> 25
<211> 20
<212> DNA
<213> Artificial
<220>
<223> laeA-AnF
<400> 25
acaactaccg ggcgactaat 20
<210> 26
<211> 20
<212> DNA
<213> Artificial
<220>
<223> laeA-AnR
<400> 26
gatctgggta aagcctgcct 20
下一篇:一种己二腈的生产方法
上一篇:一种制备泛酸内酯的方法

公众号
地址:上海浦东新区福山路388号27楼
咨询电话:18016058776
邮箱:inquires@chemdodgen.com